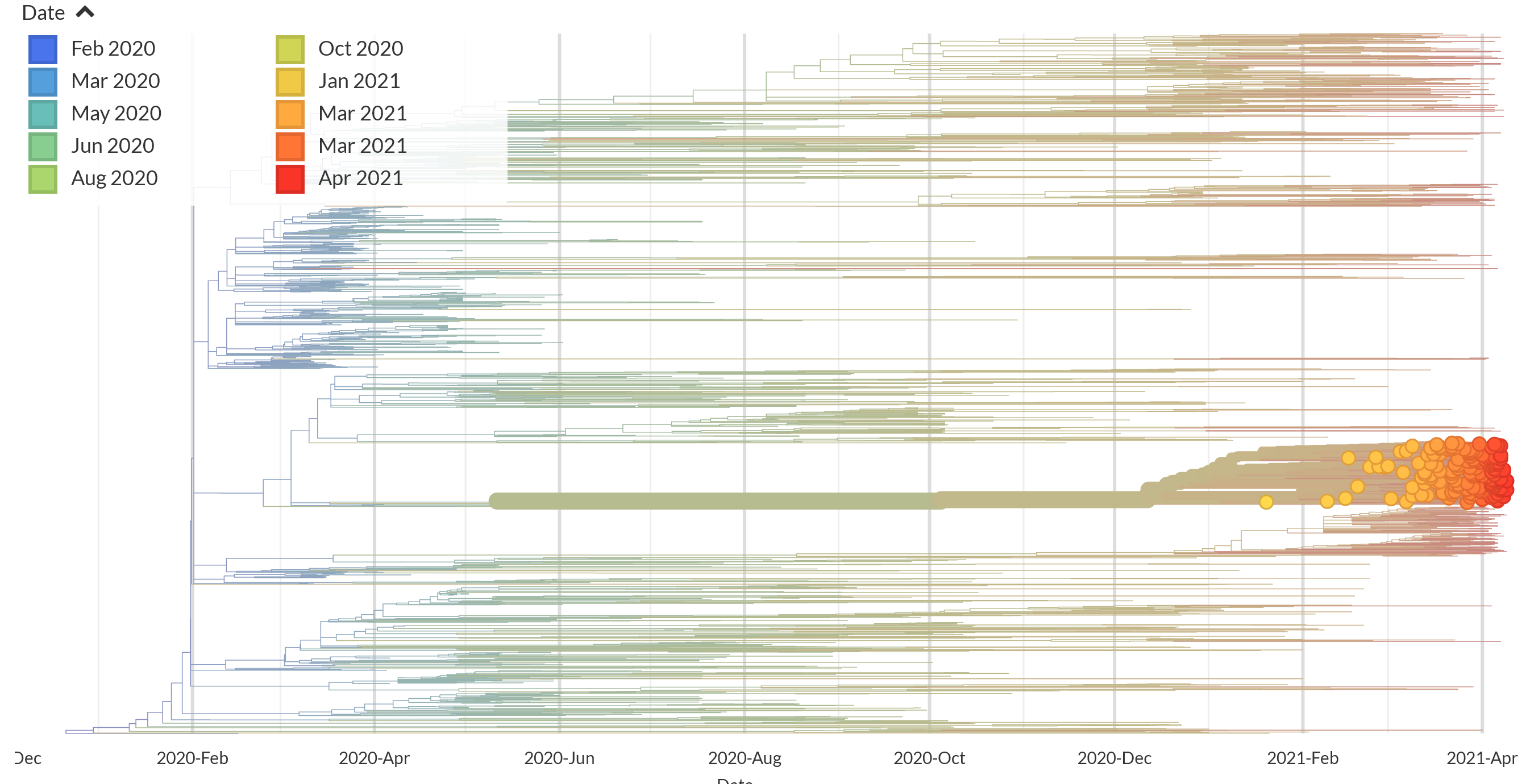
I also prepared a Nextstrain build using 1896 genomes from Chile. 161 out of 698 genomes correspond to the new C.37 lineage